Biofilm og bakteriekommunikasjon – mulige konsekvenser for oral profylakse
Forfattere
professor, dr.odont.
NFR postdoc stipendiat, dr.odont.
Institutt for oral biologi, Det odontologiske fakultet, Universitetet i Oslo
Bortsett i fra fluorider er det få kjemiske stoffer som i nevneverdig grad virker forebyggende på plakkrelaterte orale sykdommer som karies og periodontitt. En forklaring kan være at bakteriene som er involvert, organiserer seg i såkalte biofilmer. Biofilmer er komplekse tredimensjonale samfunn av bakterier. Mens mikrobiologer tradisjonelt har studert bakterier i flytende medier i laboratoriet, vet vi nå at bakterier i naturen, biofilmbakterier, uttrykker andre egenskaper enn de frittflytende. Senere tiders muligheter for å studere genekspresjon og genregulering i bakterier har gitt ny kunnskap om kommunikasjonssystemer, «quorum sensing»-systemer, som tillater bakteriene å koordinere sine genuttrykk. Vi mangler fortsatt informasjon om hvordan bakteriene regulerer sine gener for å danne dentalt plakk, såkalt oral biofilm. Mer kunnskap om disse prosessene er viktig for å utvikle nye strategier for å forebygge sykdommer forårsaket av orale biofilm-bakterier. Å interfere med bakterienes kommunikasjonssystemer kan vise seg som lovende metoder. Det er derfor en utfordring for odontologisk forskning å belyse og forstå hvordan slike kommunikasjonssystemer virker hos orale bakterier. Denne artikkelen diskuterer disse aspektene.
Ordet biofilm dukker stadig oftere opp i litteraturen, og antall publikasjoner som omhandler emnet, har økt eksponentielt siden begrepet ble tatt i bruk på slutten av syttitallet (1). Men hva er egentlig biofilm og hvilke konsekvenser har det at bakteriene velger å leve sammen i biofilm? Tradisjonelt har bakterier blitt sett på som enkle, encellede organismer, og mikrobiologer har i hovedsak studert bakterier som vokser i flytende medier eller på bakterieskåler. Dette har gitt oss nyttig kunnskap og forståelse av sammenhengen mellom bakterier og sykdom. Imidlertid har man ved denne tilnærmingen gått glipp av informasjon om viktige sider av bakterienes liv. Det viser seg nemlig at bakterier i virkeligheten ikke lever isolert som encellede organismer. Som oftest lever de festet på overflater organisert i komplekse bakteriesamfunn som vi kaller biofilm. Dentalt plakk er en typisk biofilm.
Det er interessant at Antonie van Leeuwenhoek med sitt selvkonstruerte mikroskop beskrev den orale mikroflora som en biofilm allerede på sekstenhundretallet. Han interesserte seg for plakk, «scurf», som han kalte det. Bakteriene han så i mikroskopet sitt kalte han «animalcules». I et brev fra 1683 skriver han om sine observasjoner; «The number of these animalcules in the scurf of a man is so many that I believe they exceed the number of men in a Kingdom». Videre gjorde han forsøk med vineddik som antibakterielt middel. «I took in my mouth some very strong wine-vinegar, and closing my teeth, I gargled and rinsed them very well with the vinegar. Afterwards I washed them very well with fair water, but there were an innumerable quantity of animalcules yet remaining in the scurf upon the teeth. I took a very little wine-Vinegar and mixed it with the water in which the scurf was dissolved, whereupon the animalcules died presently. From hence I conclude, that the vinegar with which I washed my teeth, killed only those animalcules which were on the outside of the scurf, but did not pass through the whole substance of it». Dette beskriver viktige egenskaper ved bakterielle biofilmer.
Bakterienes livsstil i en biofilm
En bakteriell biofilm består av bakterier festet til et underlag i en omgivende matriks. Matriksen som holder bakteriene sammen, er en blanding av polysakkarider, proteiner og DNA som stammer fra bakteriene (2). Bakterier i biofilm danner tredimensjonale strukturerte samfunn med væskekanaler for blant annet transport av næringssubstrater, avfallsprodukter og ulike signalmolekyler (3).
Bakterier i biofilm er langt mindre sensitive overfor antimikrobielle stoffer og vertens immunforsvar enn når de lever fritt i flytende medium (4). Konsentrasjonen av et stoff som dreper fritt flytende bakterier må økes, gjerne fra ti til tusen ganger for å ha tilsvarende effekt når bakteriene vokser i biofilm (4). Dette er nettopp hva van Leeuwenhoek i sin tid observerte med vineddiken han testet. Denne tilsynelatende resistensen kan forklare hvorfor mange stoffer benyttet til oral profylakse har dårligere effekt klinisk enn forventet ut i fra laboratorietester.
En forklaring på denne tilsynelatende resistensen har vært at strukturen i en biofilm hindrer diffusjon av aktive molekyler inn i dybden av biofilmen, eller at molekylene inaktiveres i overflaten av biofilmen. En annen forklaring på den tilsynelatende resistensen kan være bakterienes varierende veksthastighet i biofilmen. Det er slik at bakterienes sensitivitet er avhengig av deres veksthastighet. En bakteriekultur som vokser fort, vil være mest sensitiv overfor et hemmende stoff. Bakteriene vokser langsommere i biofilm enn i flytende medium. Derved blir bakteriene mindre sensitive.
I en biofilm dannes ulike mikromiljøer på grunn av gradienter av for eksempel næring, avfallsprodukter, pH og oksygen. Dette registrerer bakteriene og tilpasser seg de varierende forhold ved å skru av og på gener etter sine egne behov. En slik tilpasning krever regulering av et stort antall gener. Nyere forskning har vist at bakteriene på denne måten endrer egenskaper når de befinner seg i en biofilm.
Når for eksempel Streptococcus mutans vokser i biofilm, blir ca. 20 % av dens gener enten oppregulert eller nedregulert i forhold til når den lever i flytende medium (5). Enzymene som S. mutans bruker til å lage polysakkarider uttrykkes også i forskjellig grad avhengig av om biofilmen er tykk og moden eller tynn og umoden (6). Andre eksempler på endret genaktivitet i biofilm er enzymet urease i Streptococcus salivarius og sekretering av en rekke proteiner i Actinobacillus actinomycetemcomitans (7).
Biofilmdannelse i Pseudomonas aeruginosa, som forårsaker alvorlig lungebetennelse hos pasienter med cystisk fibrose, er mye studert. I biofilm sammenlignet med frittflytende bakterier og i ulike stadier av biofilmdannelsen har man funnet mellom 40 og 50 % forskjell i uttrykte gener hos denne bakterien (8).
Biofilmdannelse er en trinnvis prosess som starter med adhesjon av frittflytende bakterier til en proteindekket overflate, etterfulgt av kolonisering, koadhesjon, modning og løsning av enkelte bakterier (Fig. 1). Det er gode holdepunkter for at uttrykk av gener som er involvert i de ulike prosessene er koordinerte og velregulerte (8 – 10). Dette skjer via forskjellige signalsystemer som tillater bakteriene å kommunisere med hverandre. I bakteriene setter kommunikasjonen i gang kaskader av reaksjoner. Resultatet kan være aktivering eller hemning av transkripsjon av ulike gener.
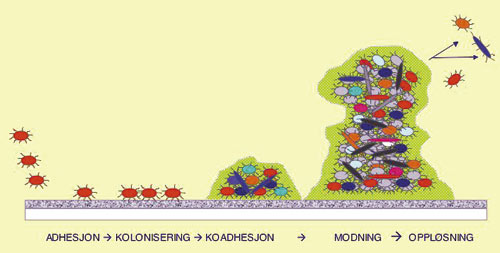
Fig. 1. De ulike trinn i biofilmdannelse.
Bakteriene kan reagere på eksterne stimuli i miljøet. I dette tilfellet kan de bruke signalmolekyler som vi ennå ikke kjenner. I andre signalsystemer er molekylene bedre kjent. Bakterietetthet er en type stimulus som kan sette i gang signalsystemer. Slike bakterie-tetthetsavhengige signalsystemer kalles «quorum sensing»-systemer. Ordet «quorum» betyr «det minste antall medlemmer av en forsamling som må være tilstede for at forsamlingen skal være beslutningsdyktig».
Bakteriene benytter ulike molekyler som quorum-signaler. Konsentrasjonen av signalmolekyl vil være en funksjon av bakterietettheten. Ikke før konsentrasjonen av signalmolekyler har nådd en viss terskelverdi, quorum, vil bakterien være i stand til å registrere og respondere på signalet. Når antall bakterier når det kritiske nivå, kan de ved hjelp av quorum-signaler koordinere sine genuttrykk (Fig. 2). For bakteriene betyr dette at de kan skru av eller på viktige gener på en koordinert måte. De kan med andre ord «trekke i flokk».
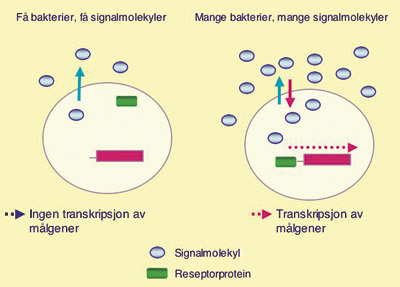
Fig. 2. Bakteriene produserer signalmolekyler som sendes ut i omgivelsene. Antall signalmolekyler vil være en funksjon av antall bakterier. Når antall bakterier har nådd en viss terskelverdi, quorum, er bakteriene i stand til å registrere signalet som setter i gang transkripsjon av gitte målgener.
Vi finner mange quorum-signalsystemer i både grampositive og gramnegative bakterier. Bakteriene benytter ulike signalmolekyler som quorum-signaler. Grampositive bakterier benytter oligopeptider som signalmolekyler, mens gramnegative bakterier benytter homoserin-laktoner. Mange quorum-molekyler er svært spesifikke. Dette innebærer at de kun kan benyttes til kommunikasjon mellom bakterier av samme spesies. Nylig avdekket man imidlertid et annet kommunikasjonssystem, autoinduser-2-systemet (AI-2). I motsetning til de fleste andre quorum-systemer som benyttes av bakteriene til intra-spesies-kommunikasjon, kan bakteriene benytte AI-2-systemet som et mer generelt, inter-spesies, kommunikasjonssystem. Enkelt kan man si at ved hjelp av det tidligere beskrevne quorum-systemet kan bakterien kommunisere med og kontrollere sine likemenn, mens AI-2 i tillegg kan benyttes til å kommunisere med andre (11).
AI-2 ble først beskrevet å stimulere bioluminiscens i marine bakterier slik som Vibrio harveyi (12). I Fig. 3 ser vi hvordan en bakteriekultur av V. harveyi skrur på genene for luminiscens og lyser med et blålig lys når tettheten av bakteriekulturen når et visst nivå. Senere studier har bekreftet at en rekke både grampositive og gramnegative bakterier har gener for å produsere AI-2-signalmolekyler (13, 14). Strukturen til AI-2 fra V. harveyi ligner ikke noen andre kjente signalmolekyler. For de fleste andre bakterier kjenner man ikke molekylstrukturen for AI-2. Man vet imidlertid at produksjonen av AI-2 avhenger av et enzym som kodes for av genet LuxS. DNA-sekvensen for dette genet ser nokså lik ut i mange bakterier (15).
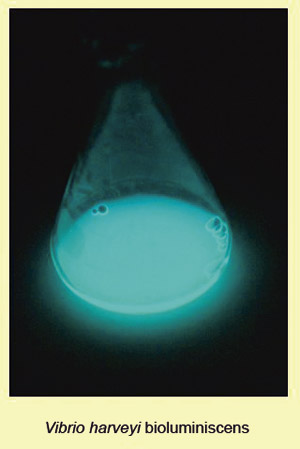
Fig. 3. Bioluminiscens, lysende blått lys, uttrykkes ved at visse gener transkriberes, «skrus på» ved en viss bakterietetthet, quorum i V. harveyi.
Kommunikasjon i plakkbiofilm
Det er rimelig å anta at det foregår livlig kommunikasjon mellom bakteriene i plakkbiofilm. I laboratoriet har man påvist evnen til kommunikasjon hos ulike orale bakterier (Tabell 1). Kommunikasjon via signalsystemer ble først vist i Streptococcus gordonii (16). Ved å inaktivere et gen som regulerer evnen til naturlig transformasjon i S. gordonii, fant man at de også mistet evnen til å danne biofilm. Naturlig transformasjon innebærer en tilstand der bakteriene er i stand til å ta opp DNA fra omgivelsene, inkorporere det nye DNA i sitt eget genom, og derved få endrete egenskaper. Man tenker seg at streptokokkene ved slik naturlig transformasjon lett kan tilpasse seg skiftende miljøforhold ved å tilegne seg egenskaper fra andre bakterier. Naturlig transformasjon i S. gordonii reguleres av et quorum-signalsystem. Signalmolekylet i dette systemet er et peptid, det såkalte kompetansestimulerende peptid (CSP) (17). Ved å inaktivere gener knyttet til kompetanseutvikling, for eksempel genet som koder for CSP i S. mutans, fant man at bakteriens evne til å lage biofilm ble sterkt redusert (18, 19). I Fig. 4 ser vi hvordan vi ved å tilsette CSP stimulerer bakteriene til å danne mer biofilm. I tillegg til å indusere naturlig transformasjon regulerer CSP også bakterienes evne til å tåle stress (20). For eksempel er dette systemet involvert i evnen S. mutans har til å tilpasse seg et surt miljø (21).
Signal- molekyl |
Bakterie |
Fenotypisk effekt på |
|---|---|---|
CSPa) |
Streptococcus mutans |
Biofilmdannelse, biofilmstruktur, naturlig transformasjon, stress og syre toleranse, celle-separasjon |
Streptococcus gordonii Streptococcus intermedius |
Biofilmdannelse, naturlig transformasjon |
|
Streptococcus mitis Streptococcus oralis Streptococcus sanguis Streptococcus crista Streptococcus anginosus Streptococcus constellatus |
Naturlig transformasjon |
|
AI-2b) |
Streptococcus gordonii |
Biofilmdannelse med Porphyromonas gingivalis, karbohydratmetabolisme |
Porphyromonas gingivalis |
Biofilmdannelse med Streptococcus gordonii, protease og hemagglutinin aktivitet |
|
Fusobacterium nucleatum |
Ukjent |
|
Prevotella intermedia |
Ukjent |
|
Actinobacillus actinomycetemcomitans |
Adaptasjon til jern-limitert miljø, leukotoksin- produksjon |
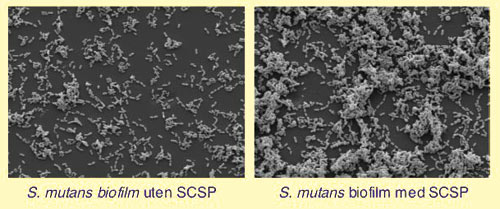
Fig. 4. Biofilmdannelse av orale streptokokker ved quorum-signalering indusert av syntetisk kompetansestimulerende peptid, SCSP.
Signaloverføring via AI-2 systemet og genet for LuxS er påvist i flere orale bakterier, for eksempel i Porphyromonas gingivalis, Prevotella intermedia, Fusobacterium nucleatum, A. actinomycetemcomitans, S. mutans (22, 23) og S. gordonii (24 – 27) (Tabell 1).
Betydningen av AI-2-signalering for biofilmdannelse i S. mutans (22, 23) og S. gordonii (24) er ikke avklart. Inaktivering av LuxS-genet synes imidlertid å påvirke S. mutans i biofilmdannelsen når den har tilgang til sukker (23). I S. gordoni, fører tilsvarende inaktivering til endret uttrykk av enzymer knyttet til karbohydrat-metabolismen (24). P. gingivalis endrer uttrykk av virulensfaktorer slik som haemagglutinin og protease-aktivitet når LuxS-genet inaktiveres (25, 26). I A. actinomycetemcomitans stimuleres leukotoksin-produksjonen av AI-2 (26). AI-2 er også involvert i bakteriens tilpasning til et miljø som har begrenset tilgang på jern (26, 27).
Siden AI-2 systemet benyttes for inter-spesies-kommunikasjon, er studier av biofilmer med flere typer bakterier særlig viktig. S. gordonii og P. gingivalis danner normalt biofilm sammen. Interessant nok ble det nylig vist at inaktivering av LuxS-genet i P. gingivalis og i S. gordonii førte til at biofilmdannelsen ble sterkt redusert (24).
Profylakse
Den senere tids utvikling og spredning av multiresistente bakteriestammer og hyppige forekomst av behandlingsresistente biofilminfeksjoner gjør at man søker andre måter å bekjempe bakterielle infeksjoner på. Tiden er inne for å gå fra bruk av antibakterielle til antipatogene stoffer. Det vil si, istedenfor å ta livet av bakteriene bør man søke å gjøre dem mindre farlige ved å regulere og begrense deres uttrykk av virulensfaktorer.
En mulig vei i fremtiden vil kunne være å interferere med bakterienes kommunikasjonssystemer. Ved hjelp av tilgjengelige gensekvensdata fra bakterielle genomer og fra det humane genom kan man identifisere aktuelle genmål som er unike for bakterier.
For terapeutiske formål vil essensielle gener for bakterienes liv representere aktuelle mål. For profylaktiske formål er det rimelig å interferere med bakterienes muligheter for å forårsake sykdom uten at balansen i normalfloraen forstyrres.
I munnhulen kan man tenke seg å benytte slike mekanismer for eksempel for å etablere en økologisk balanse som opprettholder oral helse.
Vi antar at en moden plakk-biofilm er resultatet av en serie velregulerte prosesser, hvor hver av dem kan representere et potensielt mål for biofilm-kontroll. I det følgende vil vi diskutere hvordan vi tenker at interferens med signaloverføring kan utnyttes som profylaktisk metode.
Interferens med signaloverføring
Den australske rødalgen Delisea pulchra unngår å bli kolonisert av bakterier ved at den produserer såkalte furanoner som sekundære metabolitter. Furanon har en tilsvarende struktur som quorum-signalmolekylene homoserinlaktoner. Homoserinlaktoner benyttes som quorum-signal av gramnegative bakterier. Furanoner kan syntetiseres og er vist å kunne hemme biofilmdannelse i P. aeruginosa (28). Man har også funnet andre molekyler i naturen som kan forstyrre homoserin-laktonmediert signaloverføring, for eksempel molekyler produsert av bacillus og variovorax-spesies (29, 30).
Ved å fjerne acylgruppen på homoserinlaktoner vil signaloverføringen forstyrres. Enzymet acylase I fra svin ble nylig vist å være i stand til å deacylere homoserinlaktoner fra gramnegative bakterier (31). I marine bakterier førte dette til redusert biofilmdannelse.
En annen mulighet til å interferere med signaloverføring er å benytte syntetiske signalmolekylanaloger. Det er rapportert om både agonister og antagonister (32, 33).
Staphylococcus aureus benytter tiolakton oligopeptider som signalmolekyler. Disse kan virke både som aktivatorer og inhibitorer av quorum-signaler. Naturlig forekommende signalmolekyler med små endringer kan redusere virulensen i S. aureus (34).
I odontologisk sammenheng er det interessant at det bredspektrede triclosan som benyttes i blant annet tannpastaer, hemmer syntese av forløperen for homoserinlaktoner (35). Homoserinlakton-signaler er i midlertid ikke påvist i orale bakterier (14, 25).
De kjente quorum-signalsystemene i orale bakterier inkluderer AI-2-systemet og det peptidinduserte signalsystemet (CSP). AI-2-systemet anses som et universelt quorum-system som benyttes av både grampositive og gramnegative bakterier. Derfor kan AI-2 representere et interessant mål for profylakse. Det er vist at AI-2 påvirker bindingen av P. gingivalis til S. gordonii i biofilm (24). Vi mangler fortsatt kunnskap om AI-2s rolle i plakkbiofilmer. Slik kunnskap er nødvendig for å vurdere mulighetene for profylaktisk intervensjon via dette systemet.
Å interferere med CSP-signalsystemet kan være en annen måte å bekjempe biofilmrelaterte sykdommer på. S. gordonii og S. mutans’ evne til å danne biofilm kunne tenkes redusert ved å interferere med dette systemet. Vi kjenner foreløpig ikke til om tidlige kolonisatorer som Streptococcus oralis, Streptococcus sanguis, og Streptococcus mitis, som alle produserer CSP og bruker signalet for å indusere naturlig kompetanse, også benytter signalet i biofilmdannelse. Det vil være interessant å finne ut om tilsetting av syntetisk CSP kan indusere til biofilmdannelse når det nødvendige quorum ikke er til stede. I tilfelle kunne CSP benyttes til å etablere en beskyttende biofilm som kunne forhindre kolonisering av mer virulente stammer. Vi vet at quorum-systemer og andre signalsystemer avhenger av miljøfaktorer. Det er derfor vesentlig å finne ut om de opererer, også under relevante kliniske betingelser.
Perspektiv
Dersom forebyggelse av oral sykdom skal basere seg på angrep på, eller forebyggelse av biofilmdannelse, er det vesentlig å vite om bakteriene har utviklet spesifikke mekanismer for å leve et liv i biofilm. I så fall vil mekanismene som inngår i biofilmdannelsen være konserverte og egnede mål for intervensjon. Stadiene i biofilmdannelse synes å følge tilnærmet samme mønster i ulike bakterier, men biofilm-arkitekturen og de molekylære mekanismer som er involvert, varierer. Mekanismene for biofilmdannelse i P. aeruginosa er vel karakterisert og tjener som modell for hypoteser om mekanismene i andre bakterier. Muligheten for å interferere med quorum-systemer i orale bakterier representerer en interessant mulighet for å forebygge oral sykdom i fremtiden. Imidlertid mangler vi fortsatt informasjon om de molekylære mekanismene og signalveiene som er involvert i dannelse av plakkbiofilm. Her ligger en viktig forskningsmessig utfordring.
English summary
Scheie AAa, Petersen FC.
Biofilm and bacterial communication – possible consequences for oral prophylaxis
84 – 9.
Few of the existing oral prophylactic agents have significant effects on oral diseases. The main reason is probably a neglect of the fact that the microorganisms involved organize into complex biofilm communities with features that differ from free floating cells. Recent approaches to study microbial gene expression and regulation in non-oral microorganisms have shed light on quorum sensing systems for transduction of signals that allow coordinated gene expression. Information is still lacking on the genetic regulation of dental plaque formation. A better understanding of these processes is of considerable importance for the development of novel strategies for oral disease prevention. We suggest that interfering with quorum sensing systems represents promising strategies for prevention of oral diseases. Therefore, defining the functions of such systems in oral biofilms is the challenge for future studies.
Litteratur
1. Costerton JW, Geesey GG, Cheng KJ. How bacteria stick. Sci Am 1978; 238; 86 – 95.
2. Sutherland IW. Biofilm exopolysaccharides; a strong and sticky framework. Microbiology 2001; 147; 3 – 9.
3. Costerton JW. Microbial biofilms. Annu Rev Microbiol 1995; 49; 711 – 45.
4. Davies D. Understanding biofilm resistance to antibacterial agents. Nat Rev Drug Discov 2003; 2; 114 – 22.
5. Svensäter G, Welin J, Wilkins JC, Beighton D, Hamilton IR. Protein expression by planktonic and biofilm cells of Streptococcus mutans. FEMS Microbiol Lett 2001; 205; 139 – 46.
6. Burne RA, Chen YY, Penders JE. Analysis of gene expression in Streptococcus mutans in biofilms in vitro. Adv Dent Res 1997; 11; 100 – 9.
7. Li YH, Chen YY, Burne RA. Regulation of urease gene expression by Streptococcus salivarius growing in biofilms. Environ Microbiol 2000; 2; 169 – 77.
8. Sauer K, Camper AK, Ehrlich GD, Costerton JW, Davies DG. Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm. J Bacteriol 2002; 184; 1140 – 54.
9. Davies DG, Parsek MR, Pearson JP, Iglewski BH, Costerton JW, Greenberg EP. The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 1998; 280; 295 – 8.
10. Stoodley P, Sauer K, Davies DG, Costerton JW. Biofilms as complex differentiated communities. Annu Rev Microbiol 2002; 56; 187 – 209.
11. Bassler BL. How bacteria talk to each other; regulation of gene expression by quorum sensing. Curr Opin Microbiol 1999; 2; 582 – 7.
12. Bassler BL, Wright M, Silverman MR. Multiple signaling systems controlling expression of luminescence in Vibrio harveyi; sequence and function of genes encoding a second sensory pathway. Mol Microbiol 1994; 13; 273 – 86.
13. Miller MB, Bassler BL. Quorum sensing in bacteria. Annu Rev Microbiol 2001; 55; 165 – 99.
14. Frias J, Olle E, Alsina M. Periodontal pathogens produce quorum sensing signal molecules. Infect Immun 2001; 69; 3431 – 4.
15. Schauder S, Shokat K, Surette MG, Bassler BL. The LuxS family of bacterial autoinducers; biosynthesis of a novel quorum-sensing signal molecule. Mol Microbiol 2001; 41; 463 – 76.
16. Loo CY, Corliss DA, Ganeshkumar N. Streptococcus gordonii biofilm formation; identification of genes that code for biofilm phenotypes. J Bacteriol 2000; 182; 1374 – 82.
17. Håvarstein LS, Gaustad P, Nes IF, Morrison DA. Identification of the streptococcal competence-pheromone receptor. Mol Microbiol 1996; 21; 863 – 9.
18. Bhagwat SP, Nary J, Burne RA. Effects of mutating putative two-component systems on biofilm formation by Streptococcus mutans UA159. FEMS Microbiol Lett 2001; 205; 225 – 30.
19. Yoshida A, Kuramitsu HK. Multiple Streptococcus mutans genes are involved in biofilm formation. Appl Environ Microbiol 2002; 68; 6283 – 91.
20. Yother J, Trieu-Cuot P, Klaenhammer TR, De Vos WM. Genetics of streptococci, lactococci, and enterococci; review of the sixth international conference. J Bacteriol 2002; 184; 6085 – 92.
21. Li YH, Hanna MN, Svensater G, Ellen RP, Cvitkovitch DG. Cell density modulates acid adaptation in Streptococcus mutans; implications for survival in biofilms. J Bacteriol 2001; 183; 6875 – 84.
22. Wen ZT, Burne RA. Functional genomics approach to identifying genes required for biofilm development by Streptococcus mutans. Appl Environ Microbiol 2002; 68; 1196 – 203.
23. Merritt J, Qi F, Goodman SD, Anderson MH, Shi W. Mutation of luxS affects biofilm formation in Streptococcus mutans. Infect Immun 2003; 71; 1972 – 9.
24. McNab R, Ford SK, El-Sabaeny A, Barbieri B, Cook GS, Lamont RJ. LuxS-based signaling in Streptococcus gordonii; autoinducer 2 controls carbohydrate metabolism and biofilm formation with Porphyromonas gingivalis. J Bacteriol 2003; 185; 274 – 84.
25. Burgess NA, Kirke DF, Williams P, Winzer K, Hardie KR, Aduse-Opuku J, et al. LuxS-dependent quorum sensing in Porphyromonas gingivalis modulates protease and haemagglutinin activities but is not essential for virulence. Microbiology 2002; 148; 763 – 2.
26. Fong KP, Chung WO, Lamont RJ, Demuth DR. Intra- and interspecies regulation of gene expression by Actinobacillus actinomycetemcomitans LuxS. Infect Immun 2001; 69; 7625 – 34.
27. Fong KP, Gao L, Demuth DR. luxS and arcB control aerobic growth of Actinobacillus actinomycetemcomitans under iron limitation. Infect Immun 2003; 71; 298 – 308.
28. Hentzer M, Riedel K, Rasmussen TB, Heydom A, Andersen JB, Parsek MR, et al. Inhibition of quorum sensing in
29. Dong YH, Wang LH, Xu JL, Zhang HB, Zhang XF, Zhang LH. Quenching quorum-sensing-dependent bacterial infection by an N-acyl homoserine lactonase. Nature 2001; 411; 813 – 7.
30. Leadbetter JR, Greenberg EP. Metabolism of acyl-homoserine lactone quorum-sensing signals by Variovorax paradoxus. J Bacteriol 2000; 182; 6921 – 6.
31. Xu F, Byun T, Dussen HJ, Duke KR. Degradation of N-acylhomoserine lactones, the bacterial quorum-sensing molecules, by acylase. J Biotechnol 2003; 101; 89 – 96.
32. Smith KM, Bu Y, Suga H. Induction and Inhibition of Pseudomonas aeruginosa Quorum Sensing by Synthetic Autoinducer Analogs. Chem Biol 2003; 10; 81 – 9.
33. Reverchon S, Chantegrel B, Deshayes C, Doutheau A, Cotte-Pattat N. New synthetic analogues of N-acyl homoserine lactones as agonists or antagonists of transcriptional regulators involved in bacterial quorum sensing. Bioorg Med Chem Lett 2002; 12; 1153 – 7.
34. Lyon GJ, Mayville P, Muir TW, Novick RP. Rational design of a global inhibitor of the virulence response in Staphylococcus aureus, based in part on localization of the site of inhibition to the receptor-histidine kinase, AgrC. Proc Natl Acad Sci U S A 2000; 97; 13330 – 5.
35. Hoang TT, Schweizer HP. Characterization of Pseudomonas aeruginosa enoyl-acyl carrier protein reductase (FabI); a target for the antimicrobial triclosan and its role in acylated homoserine lactone synthesis. J Bacteriol 1999; 181; 5489 – 97.
Adresse: Professor Anne Aamdal Scheie, Institutt for oral biologi, Det odontologiske fakultet, postboks 1052 Blindern, 0316 Oslo. E-post; ascheie@odont.uio.no
Artikkelen har gjennomgått ekstern faglig vurdering.