Tanden som sensor i tuggfunktionen
Denna översiktsartikel sammanfattar vad vi vet om de sinnesorgan, periodontalreceptorer, som ansvarar för att sända information till hjärnan om de krafter som träffar våra tänder.
Oralfysiologisk forskning som utförts på människa visar att periodontalreceptorerna signalerar mycket detaljerad information om position, riktning och intensitet på tugg- och bitkrafter. Denna information används av hjärnan för att reglera käkens motorik under normal funktion. Patienter med implantat, som saknar denna information, uppvisar tydliga störningar i käkens finmotoriska reglering.
Genom att öka kunskapen om hjärnans reglering av normala orala funktioner läggs grunden för en fortsatt biologisk utveckling inom oral rehabilitering. Forskningen möjliggör en utveckling av nya diagnostiska hjälpmedel som kan hjälpa klinikern att välja rätt protetisk terapi för att återskapa bästa möjliga orala funktion.
När vi till vardags använder munnen för att äta, bita och tugga behöver vi inte tänka på hur vi rör käken för att sönderdela födan så effektivt som möjligt. Vi behöver inte heller fundera över hur mycket kraft vi behöver använda för att till exempel bita av en morot. All denna exakta och mycket noggrant koordinerade motorik sker automatiskt och styrs mycket framgångsrikt av hjärnan.
För att hjärnan ska kunna styra de orala funktionerna på ett effektivt sätt krävs det att den kontinuerligt får information om vad som sker i munnen. Denna sensoriska information kommer från ett stort antal sinnesorgan som är utspridda i de orala vävnaderna (1,2).
De sinnesorgan som har huvudansvaret för att uppdatera hjärnan om de krafter som träffar tänderna är belägna i tändernas rothinnor och kallas för periodontalreceptorer (Fig. 1) (3 – 5). Det finns flera hundra periodontalreceptorer vid varje tand. När en tand belastas så kommer tandens rot att röra sig i alveolen och överföra kraft till de kollagena trådar som håller fast roten i käkbenet. Den spänning som då uppstår i de kollagena trådarna registreras av periodontalreceptorerna som skickar informationen vidare genom afferenta nervtrådar till hjärnan.
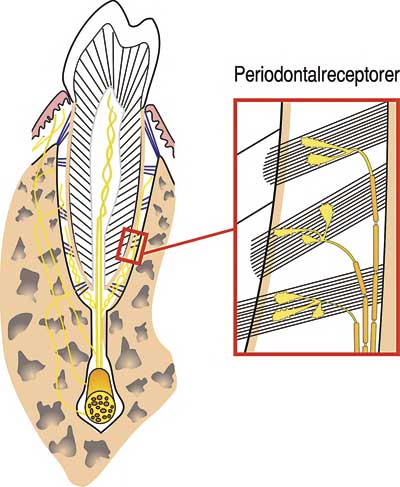
Fig. 1. Periodontalreceptorer är sinnesorgan som är specialiserade på att registrera tandbelastningar. De ligger bland de kollagena trådarna i periodontalligamentet som binder fast roten i alveolarbenet. När tanden belastas rör sig roten i alveolen. Den spänning som då uppkommer i de kollagena trådarna registreras av periodontalreceptorerna som skickar informationen vidare till hjärnan. (Modifierad från original av Marielouise Rönnmark, Umeå Universitet.)
För att vi ska kunna förstå hur hjärnan använder denna information för att styra käkmotoriken måste vi först lära oss i detalj vilken information som går in till hjärnan. Vi har inom forskargruppen under de senaste 15 åren använt mikroneurografiska nervregistreringar, direkt på människa, för att kartlägga vilken information som enskilda periodontalreceptorer skickar till hjärnan.
Nervregistreringarna utförs på friska vakna försökspersoner från nervus alveolaris inferior strax innan nerven går in i mandibularkanalen (Fig. 2A) (7,8). Denna nerv innerverar alla tänder på underkäkens ena sida men skickar också nervtrådar ut i läppen och hakan. Samtidigt som signaler registreras från enskilda periodontalreceptorer belastas tänderna för att aktivera receptorerna. Kraften på tanden mäts kontinuerligt och appliceras på små nylonkuber som fixerats centralt uppe på tanden. På detta sätt kan kraften på ett kontrollerat sätt appliceras i sex riktningar i tre dimensioner (Fig. 2B).
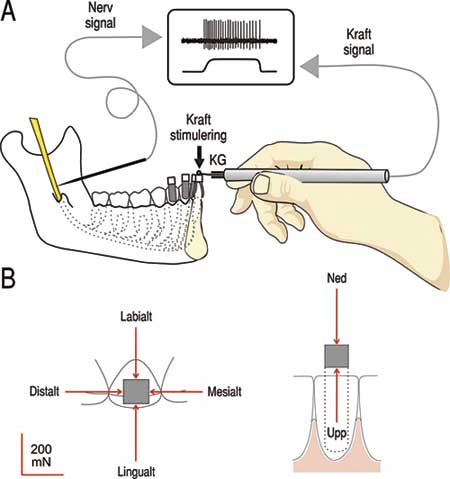
Fig. 2. Mikroneurografisk registrering av nervsignaler från periodontalreceptorer på människa. A: Medan nervsignaler registreras från enskilda nervfibrer i nervus alveolaris inferior belastas tänderna med en sond utrustad med kraftgivare (KG) så att kraften kontinuerligt kan mätas. B: Kraften appliceras i sex olika riktningar på små nylonkuber som fixerats uppe på tanden. (Modifierad från (5)).
Registreringarna från nervus alveolaris inferior visar att människans periodontalreceptorer signalerar med en stabil impulsfrekvens på ihållande tandbelastningar (statiskt svar) men ändrar också impulsfrekvensen som svar på förändringar av kraften (dynamiskt svar). Dessutom har en majoritet av receptorerna (cirka 70 %) en kontinuerlig spontan lågfrekvent signalering (cirka 5 – 10 impulser/sekund) utan att någon tand belastas. Den regelbundna «metronomliknande» signaleringen är slående och indikerar att varje nervfiber ansluter till endast en receptor i periodontalligamentet där mekanisk energi omvandlas till nervsignaler.
Signalering av belastningens läge och riktning
Periodontalreceptorer är optimerade för att registrera krafter under normal funktion, och signalerar detaljerad information till hjärnan om var på tandbågen belastningen sker och riktningen på belastningen.
Hälften av periodontalreceptorerna signalerar vid belastning av flera intilliggande tänder, men varje enskild receptor uppvisar alltid en största känslighet vid belastning av en enda tand, den «receptorbärande» tanden (9,10). Förklaringen till att en periodontalreceptor kan svara på belastning av flera tänder är att interdentala kontakter och transseptala fibrer skapar mekaniska kopplingar mellan tänderna. När en tand belastas, sprids helt enkelt kraften över till angränsande tänder där periodontalreceptorerna aktiveras. Inga forskningsresultat från människa tyder på att nervfibrerna skulle grena upp sig och innervera mer än en tand.
Vid belastning av den receptorbärande tanden svarar periodontalreceptorerna i fler än en belastningsriktning men med varierande styrka (8,10). I Fig. 3A ses ett exempel på hur en periodontalreceptor signalerar då tanden belastas med en konstant kraft (250 mN, motsvarar cirka 25g) i sex olika riktningar. Det framgår tydligt att signaleringsfrekvensen är störst då tanden trycks i distal riktning. Frekvensen ökar också då tanden trycks i labialt och mesial riktning och lyfts uppåt. Just den här receptorn visar alltså en ökad signaleringsfrekvens i fyra av den sex testade riktningarna. Typiskt för periodontalreceptorer är att visa ett ökat svar i två till fyra av de sex riktningarna.
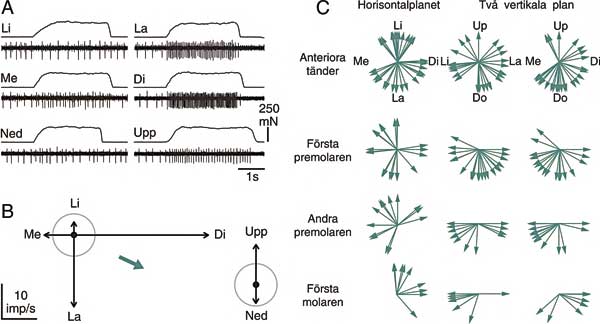
Fig. 3. A och B visar svar från en periodontalreceptor vid en framtand som belastas med en konstant kraft i sex olika riktningar. A: Exempel på kraft- och nervregistreringar i varje riktning. B: Nervsvaret i de olika riktningarna i vektorform. Riktningen på vektorerna är riktningen på kraften. Längden är den statiska signaleringen i just den riktningen. Radien på cirkeln visar spontanaktiviteten hos receptorn. Vektorer som är längre än cirkelns radie representerar en ökning av signaleringsfrekvensen. Vektorer som är kortare än radien representerar en minskning av signaleringsfrekvensen. Just den här receptorn visar en ökad signaleringsfrekvens i fyra av den sex testade riktningarna. En summering av vektorerna i varje plan (den gröna kraftiga vektorn) ger en uppskattning av den effektivaste stimuleringsriktningen («huvudriktningen») för varje receptor i det planet. C: «Huvudriktningarna» för ett antal periodontalreceptorer visas i horisontalplanet och två vertikala plan. Varje vektor representerar en receptor som svarar maximalt på belastning av en av de anteriora tänderna (incisiv eller hörntand), förstapremolaren, andrapremolaren eller första molaren. (Från (8) och (10)).
I Fig. 3B illustreras receptorns signaleringsfrekvens i de olika riktningarna som vektorer. Om svarsvektorerna i varje plan summeras så fås receptorns «huvudriktning» i det planet (grön kraftig vektor i Fig. 3B). I Fig. 3C visas huvudriktningen för alla periodontalreceptorer som vi har studerat på människa. Varje vektor illustrerar «huvudriktningen» för en periodontalreceptor i det planet. För att illustrera skillnaden i riktningskänslighet mellan receptorer vid olika tänder har receptorerna delats upp på anteriora tänder (incisiver och hörntanden), första premolaren, andra premolaren och första molaren.
Det framgår här tydligt att antalet periodontalreceptorer (vektorer) är störst för anteriora tänder och minskar posteriort i bettet. Trots att storleken på tänderna (och periodontalligamenten) generellt ökar posteriort i bettet tyder våra resultat på att antalet periodontalreceptorer minskar kraftigt posteriort. Denna slutsats får stöd av flera studier på olika djurmodeller (exempelvis (11)) och visar på betydelsen av en väl utvecklad sensorik för den främre delen av munnen. Incisiver och hörntänder är involverade i den initiala hanteringen av föda, då maten delas upp i lagom stora bitar för att sedan transporteras bakåt i munnen för att malas under tuggningen. Ibland används också framtänderna som en «tredje hand» i manipulativa uppgifter eller som ett precisionsinstrument för att bita av exempelvis en sytråd. För att dessa uppgifter ska kunna utföras på ett säkert sätt krävs en kraftig sensorisk återkoppling till hjärnan. Denna situation kan jämföras med att de finmotoriska uppgifter som utförs med handen är beroende av den kraftiga innervationen av fingertopparna.
Periodontalreceptorernas huvudriktningar i horisontalplanet är relativt väl fördelade åt alla håll för anteriora tänder och premolarer (Fig. 3C). För första molaren däremot ses en kraftig övervikt i distal och lingual riktning. Det innebär att belastningar i dessa riktningar mot första molaren ger upphov till starkare svar jämfört med andra riktningar. I vertikalplanen visar de flesta receptorerna upp en större känslighet för nedåtriktade krafter. Denna känslighet minskar dock posteriort i bettet.
Förändringen av receptorernas riktningskänslighet från alla riktningar för de anteriora tänderna till disto-lingual riktning för molaren matchar väl de funktionella krav som ställs på de olika tänderna. När de anteriora tänderna manipulerar och biter av föda då mat stoppas in i munnen belastas de i ett stort antal riktningar. Molaren utsätts däremot för en mindre variation i kraftriktningar när den maler maten med hög kraft. Under tuggning (när underkäksmolaren på arbetssidan närmar sig överkäksmolaren från en posterior och lateral position) kommer underkäksmolaren att i huvudsak belastas i distal och lingual riktning. Tack vare den utpräglade riktningskänsligheten i distal och lingual riktning är periodontalreceptorerna vid underkäksmolaren optimerade för att registrera krafter som träffar tanden under normal funktion.
Eftersom periodontalreceptorerna normalt svarar i mer än en kraftriktning kan en enskild receptor inte ge noggrann information om i vilken riktning en tand belastas. En vektoriell analys av svarsegenskaperna visar emellertid att människans periodontalreceptorer effektivt kodar belastningens riktning när hänsyn tas till informationen i en population av receptorer (8). På liknande sätt visar analyser som utförts med artificiella neuronala nätverk att en population av receptorer förmedlar detaljerad information om både kraftens riktning och vilken tand som belastas (12).
Signalering av belastningens intensitet
Periodontalreceptorer signalerar information till hjärnan om tandbelastningens storlek och hastighet. För en majoritet av receptorerna är relationen mellan tandbelastningens intensitet och nervsvarets impulsfrekvens inte linjär. Det gör att receptorerna är mycket känsliga för låga kraftnivåer.
För att undersöka hur periodontalreceptorer kodar intensiteten på tandbelastningar applicerades krafter av olika storlekar på den receptorbärande tanden i dess mest känsliga riktning (Fig. 4A) (13,14). Relationen mellan den statiska (ihållande) kraften och signaleringsfrekvensen för en grupp av receptorer vid en framtand kan ses i Fig. 4B. De flesta av människans periodontalreceptorer (cirka 80 %) uppvisar kraftigt krökta stimulus-respons relationer (se de obrutna kurvorna i Fig. 4B). Lutningen på kurvorna är brantast mellan 0 och 1 N: Det betyder att periodontalreceptorerna vid människans anteriora tänder är mest känsliga för förändringar i statiska krafter under 1 N (motsvarar cirka 100 g). Motsvarande värde för posteriora tänder är något högre (cirka 3 – 4 N). I kontrast till den branta delen av kurvan är kurvorna nästan horisontella vid högre kraftnivåer. Receptorerna satureras (mättas) över gränsen och behåller sin maximala signaleringsfrekvens oberoende av om kraften ökar. Vid höga tandbelastningar signalerar alltså de flesta periodontalreceptorer information till hjärnan om att tanden belastas men de ger mycket lite eller ingen information om storleken på belastningen.
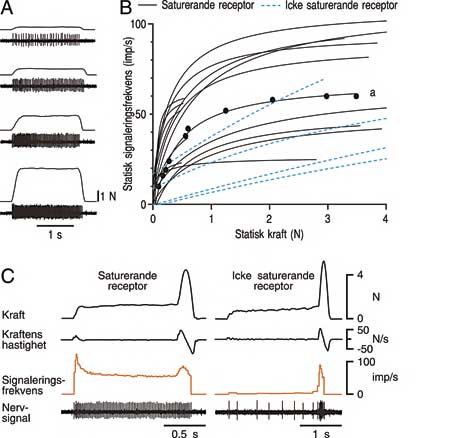
Fig. 4. Svar från periodontalreceptorer då krafter av olika storlekar appliceras på en framtand i den mest känsliga riktningen. A: Exempel på kraftstimuleringar och nervregistreringar från en periodontalreceptor som belastas med fyra olika krafter. B: Stimulus-responsrelationer för 19 stycken periodontalreceptorer. De anpassade kurvorna definieras av funktionen F/(F+c), där F är den applicerade kraften och c är en konstant som bestäms för varje receptor. Obrutna kurvor illustrerar saturerande receptorer medan blå streckade kurvor illustrerar icke saturerande receptorer. Kurvan som är markerad med a är den receptor som visas i A. C: Svar från en saturerande och en icke saturerande receptor på en snabb kraftökning som utlöses efter att tanden har belastats med en konstant kraft på cirka1 N. Notera den stora skillnaden i signaleringsfrekvensen mellan de två receptorerna. (Från (13)).
Periodontalreceptorer registrerar inte bara statiska krafter. De är också känsliga för snabba förändringar i tandbelastningen, så kallad dynamisk känslighet. Receptorernas dynamiska känslighet minskar parallellt med att känsligheten för statiska krafter minskar vid ökad belastning. Detta illustreras för en saturerande receptor till vänster i Fig. 4C. Efter att en tand belastats i cirka 1,5 sekunder med en ihållande kraft på cirka1 N ökas kraften snabbt till cirka 4 N. Receptorns svar på den snabba kraftökningen i slutet är förvånande liten. Trots att hastigheten på kraftökningen är lägre vid den initiala tandkontakten jämfört med den snabba kraftökningen i slutet är receptorns svar störst vid den initiala tandkontakten. Det beror på att saturerande periodontalreceptorer blir mindre känsliga både för storleken på statiska krafter och för hastigheten på snabba kraftförändringar i takt med att tandbelastningen ökar.
I motsatts till majoriteten av periodontalreceptorerna som saturerar vid mycket låga kraftnivåer finns det en minoritet (cirka 20 %) som visar nästan linjära stimulus-responsrelationer (se de streckade blå kurvorna i Fig. 4B). Dessa icke saturerande receptorer har kapacitet att signalera information om förändringar i statiska krafter både på låga och höga kraftnivåer. Precis som för de saturerande receptorerna följer den dynamiska känsligheten den statiska känsligheten. Det innebär att känsligheten för snabba kraftförändringar kvarstår vid högre kraftnivåer. Detta illustreras för en icke saturerande receptor till höger i Fig. 4C. Receptorsvaret domineras av det kraftiga svaret på den slutliga snabba och höga kraftökningen. Notera den stora skillnaden i svarsmönster mellan den icke saturerande receptorn (till höger) och den saturerande receptorn (till vänster) på liknande belastningar.
Signalering under funktion
Periodontalreceptorer med olika funktionella egenskaper är specialiserade på att samla in information om specifika faser under normal bit- och tuggfunktion. De flesta periodontalreceptorer svarar kraftigt på den initiala kontakten med föda, signalerar kontinuerligt information om belastningen då föda manipuleras mellan tänderna men ger lite eller ingen information om kraftnivån vid höga bit- och tuggkrafter.
Två kriterier måste ha uppfyllts för att man ska kunna säga att ett sinnesorgan som periodontalreceptorn medverkar i regleringen av ett motoriskt beteende. 1) Receptorn måste signalera värdefull information under specifika faser av det motoriska beteendet, och 2) det måste finnas bevis för att informationen verkligen används av hjärnan för att reglera beteendet. Periodontalreceptorerna uppfyller båda dessa kriterier när det gäller manipulation av föda mellan framtänderna.
Av förklarliga skäl är det inte lätt att registrera nervaktivitet från enskilda periodontalreceptorer på människa under bitning eller tuggning. För att komma runt detta problem har vi utvecklat en matematisk modell för receptorernas kodning av tandbelastningars styrka. Den innehåller en icke-linjär överföringsfunktion från kraft till nervimpulsfrekvens och uttryck för receptorernas statiska och dynamiska känslighet. Modellen kan med mycket stor noggrannhet förutsäga enskilda receptorers impulssvar vid olika kraftprofiler.
För att kunna simulera periodontalreceptorernas svar under normal funktion måste vi först registrera de krafter som utvecklas mellan tänderna under det motoriska beteendet. Vi valde en mycket enkel motorisk uppgift – att hålla och bita av föda med framtänderna (15). En bit mat (jordnöt eller en bit kaka) placerades på en liten platta utrustad med en kraftgivare (Fig. 5A). Precis som periodontalreceptorerna registrerar kraftgivaren kontinuerligt de krafter som tänderna utsätts för under bituppgiften. Kraften som träffar tänderna under en avbitning av en jordnöt (då försökspersonen först håller och sedan biter av jordnöten) kan ses överst i Fig. 5B och C. Under varje kraftprofil ses de simulerade svaren från två periodontalreceptorer – en saturerande receptor och en icke saturerande receptor.
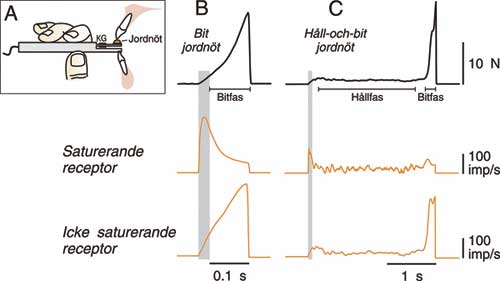
Fig. 5. Simulerade svar från periodontalreceptorer på kraftprofiler som registrerats under en bituppgift och en håll-och-bituppgift med en jordnöt. A: Utrustning som används för att mäta krafterna som utvecklas mellan tänderna. Jordnöten placeras på en aluminiumplatta utrustad med en kraftgivare (KG) för kontinuerlig kraftregistrering. Försökspersonen instrueras att placera plattan med jordnöten mellan två framtänder och bita av den, eller hålla jordnöten mellan tänderna och sen bita av den. B och C: Nedanför kraftprofilerna som registrerats under en bituppgift (B) och en håll-och-bituppgift (C) ses de simulerade receptorsvaren från en saturerande receptor och en icke saturerande receptor. Notera att de två receptortyperna signalerar helt olika information om kraften som utvecklas under den motoriska uppgiften. (Från (15) och (16)).
Modellen visar att den saturerande periodontalreceptorn svarar distinkt på de små krafter som uppstår under den initiala kontakten med jordnöten (skuggade områden i Fig. 5B och C). Receptorn signalerar kontinuerligt under det att försökspersonen håller jordnöten mellan tänderna (hållfas) (16). Under dessa låga krafter signalerar receptorn viktig information om kraftnivån och om förändringar i kraften. Däremot svarar receptorn mycket dåligt på de höga krafter som krävs för att bita av jordnöten. Trots att kraften under bitfasen ökar kontinuerligt tills jordnöten spricker minskar samtidigt receptorns signalering. I motsats till den saturerande receptorn kommer den icke saturerande receptorn att signalera information om kraftnivån under hela bitfasen.
Liknande modeller har konstruerats för periodontalreceptorer vid posteriora tänder. De har använts för att simulera receptorernas svar under tuggning (14). Precis som för de anteriora periodontalreceptorerna svarar en majoritet av de posteriora receptorerna kraftigt på den initiala kontakten med födan men ger lite eller ingen information om kraftnivån vid de höga tuggkrafter som används för att mala sönder maten.
Reglering av käkmotorik
Hur kan vi veta att periodontalreceptorerna verkligen är involverade i kontrollen av oral motorik?
Eftersom majoriteten av framtändernas periodontalreceptorer är mest känsliga för tandbelastningar under 1 N och kontinuerligt signalerar information om belastningen under hållfasen under en håll- och bituppgift är det rimligt att förvänta sig att signalerna från dessa receptorer är av betydelse för regleringen av de låga krafter som används under hållfasen. I Fig. 6A ses exempel på fem stycken håll-och-bitkraftprofiler som lagts på varandra och i Fig. 6B ses ett histogram på hållkrafterna från ett stort antal tester på flera försökspersoner (15). Trots att försökspersonerna inte fått någon information om vilken kraftnivå de skulle använda för att hålla jordnöten mellan tänderna använder de spontant en hållkraft som vanligen ligger under 1 N. Kurvorna i Fig. 6C visar hur känsligheten hos framtändernas periodontalreceptorer minskar när kraften ökar. Försökspersonerna väljer automatiskt en hållkraft som är lagom stor för att ha en stabil kontakt med jordnöten (medelvärde cirka 0,6 N) men undviker högre hållkrafter då känsligheten hos receptorerna går ner. Det verkar alltså som att försökspersonerna omedvetet använder informationen från periodontalreceptorerna för att välja en hållkraft som ger en god kontroll av jordnöten. Om denna hypotes stämmer bör det motoriska beteendet störas om signalerna från receptorerna tas bort. Detta låter sig enkelt testas genom att man bedövar tänderna och återupprepar håll-och-bituppgiften. Från kraftprofilerna i Fig. 6B framgår att bedövningen leder till en påtaglig störning av kraftregleringen under hållfasen medan bitfasen är i det närmaste opåverkad (se också histogrammet i Fig. 6C). Under bedövning är hållkraften cirka 4 gånger högre jämfört med normal parodontal känslighet (se diagrammet till vänster i Fig. 6D).
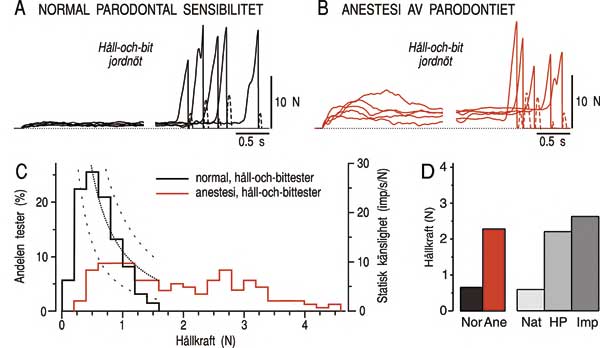
Fig. 6. Periodontalreceptorer är viktiga för regleringen av kraftnivån under hållfasen. A och B: Exempel på kraftprofiler (fem stycken) som registrerats under en håll-och-bit-uppgift med en jordnöt då försökspersonen haft normal parodontal känsel (A) och då parodontiet varit bedövat (B). Notera att betydligt högre krafter användes för att hålla jordnöten mellan tänderna då signalerna från periodontalreceptorerna blockerats med bedövning. C: Histogram som visar hållkrafter från ett stort antal tester från flera försökspersoner under normala förhållanden och under anestesi. De streckade kurvorna visar hur känsligheten hos periodontalreceptorerna vid framtänderna sjunker när kraften ökar. Kurvorna representerar första derivatan av medelvärdet (+/- 1 SD) av stimulus-respons kurvorna för de 19 periodontalreceptorerna i Fig. 4 B (15). D: De två vänstra staplarna visar hållkraftens medelvärde för en grupp försökspersoner under normal sensibilitet (Nor) och under anestesi av tänderna (Ane) (15). De tre högra staplarna visar hållkraftens medelvärde för tre olika grupper: Försökspersoner med naturliga tänder (Nat), helproteser i bägge käkarna (HP) och implantatbroar i bägge käkarna (Imp) (17). Notera likheten mellan försökspersoner som är bedövade (Ane) och försökspersoner som saknar periodontalreceptorer (HP, Imp).
Intressant nog har det också visat sig att personer som helt saknar information från periodontalreceptor får en störd reglering av hållfasen (17). Personerna, som har helproteser eller implantatbroar i bägge käkarna, använder hållkrafter som ligger på samma nivå som vid bedövning (se diagrammet till höger i Fig. 6D). Andra mekanoreceptorer i munhålan eller i käkmusklerna kan inte på ett fullgott sätt ersätta periodontalreceptorerna i denna kontroll.
Signaler från periodontalreceptorer är nödvändiga för en normal reglering av låga krafter mellan käkarna, exempelvis då föda manipuleras och positioneras för att senare sönderdelas under tuggning.
Håll-och-bittestet är normalt mycket enkelt att utföra. Med normal parodontal sensibilitet kan man lätt placera tänderna rätt på jordnöten och rikta in bitkraften åt rätt håll så att jordnöten knäcks under bitfasen. Om det däremot saknas information från periodontalreceptorerna (vid bedövning eller patienter med enbart implantat) misslyckas försökspersonen oftare med att knäcka jordnöten. Detta beror på att periodontalreceptorerna inte längre signalerar information om läget och riktningen på bitkraften. Hjärnan kan inte längre finjustera käkmusklerna så att bitkraftvektorn riktas rakt igenom jordnöten. Istället för att knäckas glider jordnöten iväg (5, 15).
Information från periodontalreceptorer är viktig för att bitkraftens position och riktning ska kunna anpassas till födans placering på tandbågen. Patienter som saknar naturliga tänder och istället har implantat visar tydliga störningar i käkens finmotoriska reglering.
Hur använder hjärnan sensorisk information för att reglera käkmotorik?
Det har tidigare föreslagits att signaler från periodontalreceptorer används av hjärnan för att öka tuggkrafterna då vi tuggar på något hårt (positiv feed-back) men att de också kan användas för att «stänga av» musklerna då de höga krafterna riskerar att skada tänderna (negativ feed-back) (18). De icke saturerande periodontalreceptorerna har förmågan att signalera information om höga bit- och tuggkrafter och kan därigenom medverka i en sådan kontroll.
Motorisk reglering genom feed-back av sensorisk information är av stor betydelse för kontrollen av statiska positioner (exempelvis hållfasen under en håll-och-bituppgift) eller långsamma motoriska förlopp. Vid regleringen av snabba motoriska förlopp (exempelvis avbitning eller tuggning) tillskrivs i dag för feed-forwardmekanismer en större betydelse (exempelvis (19)). Här samlar man tidigt in sensorisk information som används för att uppdatera ett motoriskt program innan beteendet uttrycks. På detta sätt kan snabb motorik anpassas till rådande förhållanden utan att behöva «vänta» på sensorisk återkoppling.
Den höga känsligheten för låga kontaktkrafter för de saturerande periodontalreceptorerna gör att dessa receptorer signalerar kraftfullt vid den initiala kontakten med födan vid en avbitning eller en tuggcykel (Fig. 5B och C). I detta ögonblick signaleras information om position och riktning på kraften, samt troligen också information om födans mekaniska egenskaper. Denna information används av hjärnan för att uppdatera de motoriska program som används för att styra de höga och snabba krafter som utvecklas vid avbitning och tuggning. På så sätt kan käkmusklernas aktivitet snabbt anpassas till födans hårdhet och läge på tandbågen så att den motoriska uppgiften kan utföras på ett säkert och effektivt sätt (5).
Periodontalreceptorerna är sannolikt också viktiga för regleringen av de höga krafter som utvecklas mellan tänderna då vi biter och tuggar.
Genom att öka kunskapen om hjärnans reglering av normala orala funktioner läggs grunden för en fortsatt biologisk utveckling inom oral rehabilitering. Forskningen skapar förutsättningar för att förstå hur olika terapiinsatser påverkar den orala funktionen. Den möjliggör även en utveckling av nya diagnostiska hjälpmedel som kan hjälpa klinikern att välja rätt protetisk terapi för att återskapa bästa möjliga orala funktion.
English summary
The tooth as a sensor in the masticatory system
Teeth are equipped with periodontal mechanoreceptors that signal information to the brain about tooth loads. In this review, the functional properties of human periodontal receptors are presented along with a discussion about their propable functional role in the control of human mastication. Microneurographic recordings from single nerve fibers in the inferior alveolar nerve reveal that human periodontal receptors adapt slowly to maintained tooth loads. Populations of periodontal receptors encode information about which teeth are loaded and the direction of forces applied to individual teeth. Most receptors exhibit a markedly curved relationship between discharge rate and force amplitude, featuring the highest sensitivity to changes in tooth load at very low force levels. Accordingly, periodontal receptors efficiently encode tooth load when subjects contact and gently manipulate food by the teeth. In contrast, only a minority of the receptors encodes the rapid and strong force increase generated when biting and chewing on food. It is demonstrated that periodontal receptors are particularly important for the control of jaw actions associated with intraoral manipulation of food. Thus, patients who lack information from periodontal receptors show an impaired fine motor control of the mandible.
Fakta
Mikroneurografi
Registreringsteknik som möjliggör att man, direkt på en vaken människa, kan «tjuvlyssna» på de aktionspotentialer som signaleras från ett enskilt sinnesorgan till hjärnan. Metoden som vanligen används för att studera sensorik i armar och ben utvecklades av två svenska forskare på 1960-talet (6).
Fakta
Sensori – motorisk reglering
Sensorisk information kan användas av hjärnan på två principiellt olika sätt för att styra motorik.
Feed-back
Sensorisk information samlas in under hela den motoriska uppgiften och används kontinuerligt av hjärnan för att korrigera motoriken. Används för att reglera statiska positioner eller långsam motorik som exempelvis när vi står upp och håller balansen.
Feed-forward
Sensorisk information används av hjärnan för att uppdatera motoriska program som sedan styr motoriken. Används för att reglera snabb motorik som exempelvis då vi fångar en boll som kastas mot oss.
Referenser
1. Trulsson M, Essick GK. Mechanosensation. In: Clinical oral physiology. Miles T, Nauntofte B, Svensson P, editors. Copenhagen: Quintessence; 2004. p. 165 – 97.
2. Trulsson M, Johansson RS. Orofacial mechanoreceptors in humans: encoding characteristics and responses during natural orofacial behaviors. Behav Brain Res 2002; 135: 27 – 33.
3. Hannam AG. The innervation of the periodontal ligament. In: Berkovitz B, Moxham B, Newman H, editors. The periodontal ligament in health and disease. Oxford: Pergamon Press; 1982. p. 173 – 96.
4. Linden RWA. Periodontal mechanoreceptors and their functions. In: Taylor A, editor. Neurophysiology of the jaws and teeth. New York: Macmillan; 1990. p. 52 – 88.
5. Trulsson M, Johansson RS. Encoding of tooth loads by human periodontal afferents and their role in jaw motor control. Prog Neurobiol 1996; 49: 267 – 84.
6. Vallbo ÅB, Hagbarth KE. Activity from skin mechanoreceptors recorded percutaneously in awake human subjects. Exp Neurol 1968; 21: 270 – 89.
7. Johansson RS, Olsson KÅ. Micro-electrode recordings from human oral mechanoreceptors. Brain Res 1976; 118: 307 – 11.
8. Trulsson M, Johansson RS, Olsson KÅ. Directional sensitivity of human periodontal mechanoreceptive afferents to forces applied to the teeth. J Physiol 1992; 447: 373 – 89.
9. Trulsson M. Multiple-tooth receptive fields of single human periodontal mechanoreceptive afferents. J Neurophysiol 1993; 69: 474 – 81.
10. Johnsen SE, Trulsson M. Receptive field properties of human periodontal afferents responding to loading of premolar and molar teeth. J Neurophysiol 2003; 89: 1478 – 87.
11. Byers MR, Dong WK. Comparison of trigeminal receptor location and structure in the periodontal ligament of different types of teeth from the rat, cat, and monkey. J Comp Neurol 1989; 279: 117 – 27.
12. Edin BB, Trulsson M. Neural network analysis of the information content in population responses from human periodontal receptors. Proc SPIE 1992; 1710: 257 – 66.
13. Trulsson M, Johansson RS. Encoding of amplitude and rate of forces applied to the teeth by human periodontal mechanoreceptive afferents. J Neurophysiol 1994; 72: 1734 – 44.
14. Johnsen SE, Trulsson M. Encoding of amplitude and rate of tooth loads by human periodontal afferents at premolar and molar teeth. J Neurophysiol 2005; 93: 1889 – 97.
15. Trulsson M, Johansson RS. Forces applied by the incisors and roles of periodontal afferents during food-holding and -biting tasks. Exp Brain Res 1996; 107: 486 – 96.
16. Trulsson M, Johansson RS. Human periodontal mechanoreceptors: encoding of force and role in control of jaw actions. In: Morimoto T, Matsuya T, Takada K, editors. Brain and oral functions, oral motor function and dysfunction. Amsterdam: Elsevier Science; 1995. p. 155 – 63.
17 .Trulsson M, Gunne HSJ. Food-holding and -biting behavior in human subjects lacking periodontal receptors. J Dent Res 1998; 77: 574 – 82.
18. Lund JP. Mastication and its control by the brain stem. Crit Rev Oral Biol Med 1991; 2: 33 – 64.
19. Johansson RS. Dynamic use of tactile afferent signals in control of dexterous manipulation. Adv Exp Med Biol 2002; 508: 397 – 410.
Korrespondens: Mats Trulsson, Odontologiska Institutionen, Karolinska Institutet, Box 4064, 141 04 Huddinge, Sverige. E-mail: mats.trulsson@ki.se
Referensgranskad artikel.
Artikkelen er fagfellevurdert.
Artikkelen siteres som:
Trulsson M. Tanden som sensor i tuggfunktionen. Nor Tannlegeforen Tid. 2006;116:6–13. doi:10.56373/2006-1-4