Den parodontale infektion
De parodontale sygdommes ætiologi har været til debat i hele det 20. århundrede. Parodontitis har været opfattet både som en endogen, non-specifik sygdom og som en eksogen, specifik sygdom. Nu samler interessen sig om den økologiske plakhypotese, hvor ophobning af patogene bakteriearter i den fastboende flora kan udløse parodontal destruktion. Hvorvidt sygdom udvikles, afgøres af balancen mellem mikrofloraen og værtens immunforsvar.
Den parodontale infektion har været genstand for vidt forskellige opfattelser, lige fra en non-specifik til en monospecifik infektion. Artiklen gennemgår grundlaget og konsekvenserne, som leder frem til nutidens dominerende opfattelse, den økologiske plakhypotese. Denne beskriver hovedparten af de parodontale sygdomme som endogene infektioner, forårsaget af den fastboende flora. Styrkeforholdet mellem mikroorganismernes virulens og antal i forhold til immunforsvarets kompetence bliver afgørende for hvorvidt der sker parodontal nedbrydning. Betydningen af de seneste års molekylærbiologiske landvindinger for de parodontale sygdommes ætiologi bliver diskuteret.
Den non-specifikke hypotese
Parodontitis som en non-specifik sygdom (Tabel 1) var den fremherskende hypotese i 1950’erne og 1960’erne. Gruppen omkring Waerhaug påviste en epidemiologisk sammenhæng mellem mængden af plak og graden af parodontal destruktion (1,2). Disse fund var vel til dels en funktion af at der i befolkningen ikke var så stor opmærksomhed på tandbørstning som middel til parodontal sundhed. Imidlertid blev hypotesen yderligere støttet af at parodontitis kunne induceres på beagle hunde ved at ligere tænderne så der ophobedes plak (3), og af at stort set alle testede bakteriearter kunne fremkalde parodontitis på forsøgsdyr. Der var altså god grund til at antage at alle mikroorganismer i plakken bidrog til virulensen, og at den samlede plakmængde var dét der afgjorde patogeniteten. Daglig, total plakkontrol og kirurgisk eliminering af pocherne blev det foretrukne behandlingsprincip.
Parodontitis’ hypotese |
Mikrobiel årsag |
Forebyggelse og behandling |
Hypotesen giver ikke forklaring på |
|---|---|---|---|
Non-specifik |
Hele plakken |
Total plakkontrol |
Uens distribution af pocher i tandsættet |
Specifik |
Specifikke bakterier |
Fjernelse af årsag med antibiotica |
Betydning af plak uden specifikke bakterie |
Betydning af persisterende dybe pocher uden den specifikke bakterie |
|||
Økologisk |
Patogene konsortier samt deres antal og aktivitet i den fastboende flora |
Total plakkontrol for at forhindre opvækst af patogene bakterier |
|
Plakophobning er forudsætning for patogene bakteriers vækst |
If. den non-specifikke plakhypotese burde sites med samme plakbelastning over tid udvikle den samme grad af parodontal destruktion. Dette er imidlertid langt fra tilfældet. På trods af samme plakbelastning ses i størsteparten af de parodontale sygdomstilfælde en varierende grad af parodontal destruktion, spændende fra hurtig over langsom til ingen påviselig nedbrydning over lang tid (4).
Det varierede nedbrydningsmønster antyder at der er tale om forskellig grad af patogenitet i plakken, hvilket nemmest skal søges forklaret i en forskellig plaksammensætning. Forbedrede metoder til dyrkning af den subgingivale mikroflora slog igennem i 1970’erne og 1980’erne, hvor et væld af nye arter blev opdaget. Fokus var på gruppen af gramnegative stave, der var mere fremtrædende i den subgingivale end i den supragingivale flora. Forsøg på dyr under standardiserede betingelser viste forskelle i de testede bakteriearters evne til at fremkalde vævsdestruktion, og dette korrelerede ofte med forskelle i relevante virulensfaktorer mellem arterne. Herfra var det snublende nært at forestille sig en egentlig specificitet i de parodontale sygdommes ætiologi.
Den specifikke hypotese
Især i 1980’erne og 1990’erne var den specifikke hypotese fremherskende (Tabel 1). Baggrunden for hypotesens succes var en ensidig fokuseren på en række gramnegative stave. På trods af eller måske netop pga. en stigende erkendelse af mikrobiel kompleksitet i de parodontale pocher, valgte mange at fokusere på nogle få, udvalgte, formodet patogene arter, deriblandt Actinobacillus actinomycetemcomitans, Porphyromonas gingivalis og Prevotella intermedia (5). Forekomsten og mængden af disse få arter blev sammenlignet mellem syge og sunde sites i et utal af undersøgelser. Dette bekræftede de pågældende arters tilstedeværelse ved mangesyge sites og fravær ved sunde sites. I konsekvens af hypotesens opfattelse af en specifik årsag blev behandlingsmålet eliminering af den eller de specifikke mikroorganismer.Behandlingsstudier havde succes med antibiotikabehandling med tilsyneladende stop for yderligere parodontal destruktion samt eliminering af disse gramnegative stave. Den specifikke hypoteses succes var dog i høj grad en funktion af den ensidige fokuseren på få arter og negligering af de øvrige mikroorganismer. Der anvendtes bredspektrede antibiotika med virkning på hele floraen, men alligevel undersøgte man kun effekten på de få udvalgte arter. Sideløbende blev der publiceret andre undersøgelser specielt fra grupperne omkring Moore fra Virginia Polytechnic Institute og Socransky fra Forsyth Dental Center i Boston, som viste at mange arter kunne isoleres fra den subgingivale flora og med større forekomst her end supragingivalt eller ved gingivitis og sunde sites (6, 7). Endvidere spillede detektionsniveauet en rolle. Ved dyrkning undgår mikroorganismer med meget lav forekomst, som fx de gramnegative stave ved sunde forhold, ofte detektion. Molekylærbiologiske undersøgelser som ikke kræver dyrkning, fx påvisning af artsspecifikt rDNA, har vist at såkaldte parodontitis-bakterier kan detekteres under sunde forhold, men i så lille mængde at de ikke vil opdages ved dyrkning og heller ikke have en patogenetisk betydning.
Den økologiske plakhypotese
Den økologiske plakhypotese (Tabel 1) har erstattet den specifikke og den non-specifikke sygdomsmodel og er i dag den fremherskende (8). Plakophobning og gingivitis er ledsaget af et karakteristisk skift i mikrofloraens sammensætning fra en overvejende fakultativ og grampositiv flora ved sunde forhold til en tiltagende anaerob og gramnegativ flora. Plakophobningen resulterer i anaerobe forhold i de dybere lag af plakken, og gingivitis er karakteriseret ved inflammation med ekssudation af det næringsrige gingivalekssudat. Det giver netop anaerobe og ernæringsmæssigt krævende gramnegative bakterier mulighed for at øge deres andel af floraen. Heriblandt findes også de mest patogene medlemmer af mundfloraen. Den økologiske plakhypotese anerkender altså at visse bakterier i den fastboende flora er mere patogene end andre. Men det er plakophobningen der er forudsætningen for at disse arter kan vokse frem til et skadeligt niveau. Derfor er daglig, total plakkontrol vigtig. Eliminering eller regelmæssig scaling af dybe pocher er nødvendig for at forhindre en patogen sammensætning af floraen i at udvikle sig.
Dét fokus som undersøgelser fra 1980’erne og 1990’erne gav til bakteriearter som A. actinomycetemcomitans, Pr. intermedia og P. gingivalis (5) er formentlig berettiget. Der er blot ikke for alle patienter en entydig sammenhæng mellem nogle få arter og parodontitis (Fig. 1). Desuden kan de samme arter isoleres fra pocher med sygdomsaktivitet og pocher uden sygdomsaktivitet (6,7). Derfor er den overvejende opfattelse at de parodontale sygdomme er polymikrobielle sygdomme, hvor skiftende konsortier af mikroorganismer forårsager nedbrydning. I visse pocher opstår en mikrobiel sammensætning der overskrider tærsklen for et harmonisk forhold til værten, hvorefter destruktion af parodontiet bliver resultatet. Da den økologiske plakhypotese anerkender at visse bakterier er mere patogene end andre i den fastboende flora, kan destruktionsgraden variere i et tandsæt på trods af samme plakbelastning.
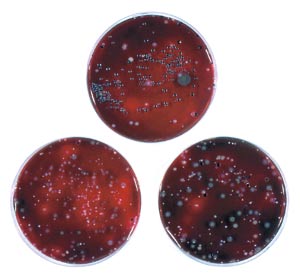
Fig. 1. Tre prøver af subgingival plak udsået på blodagarplader. Bemærk forskellen i forekomst af sort-pigmenterede kolonier. Disse kan repræsentere Porphyromonas gingivalis og Prevotella-arter.
Den parodontale mikrofloras kompleksitet
Forbedrede dyrknings- og identifikationsmetoder har øget antallet af kendte mikrobielle arter i den subgingivale plak fra ca. 50 til ca. 400 over de seneste 25 år. Helt nye undersøgelser har vha. molekylærbiologiske teknikker afsløret en hidtil uset kompleksitet i den subgingivale flora. DNA oprenses fra en plakprøve, og derefter opformeres genet der koder for 16S ribosomalt RNA, og som findes i alle bakterier, vha. den såkaldte PCR-metode (polymerase chain reaction). Man står derefter med en blanding af rDNA fra alle bakterierne i prøven. De enkelte opformerede DNA-fragmenter overføres til en Escherichia coli-bakterie vha. såkaldt kloning i et plasmid. E. coli-bakterien dyrkes op i stor mængde, så fragmentet fra bakterien kan oprenses og sekvensen bestemmes. De fremkomne sekvenser sammenlignes med de hundredetusinder af sekvenser fra kendte dyrkbare bakterier, der efterhånden er deponerede i internationale databaser. Sådanne undersøgelser har vist at kun 50 – 60 % af floraen fra subgingival plak tilhører kendte bakteriearter (9,10). Resten stammer fra ukendte arter, hvoraf størsteparten endnu ikke er dyrkede. En del af disse arter vil formentlig forekomme i meget lille antal, som gør dem irrelevante i parodontitis-patogenetisk sammenhæng, men der vil sikkert også blandt de mange ikke-dyrkbare arter være nogle der både forekommer i stort antal og har en betydelig virulens.
Hertil kommer at opmærksomheden for nylig er blevet rettet mod virus som en mulig ætiologisk faktor i parodontitis-patogenesen. Det er meget tidskrævende at dyrke virus, og ofte unddrager de sig dyrkning fordi de er til stede i meget små mængder. Vha. af PCR og såkaldte «primere», der er specifikke for en virusart, kan DNA fra bestemte virus opformeres og DNA-sekvensen bestemmes. Epstein-Barr virus og humant cytomegalovirus findes hyppigere ved parodontitis end ved gingivitis og parodontalt sunde sites (11). Pocher med disse virus indeholder samtidig forøget niveau af parodontitis-bakterier som A. actinomycetemcomitans, P. gingivalis, Bacteroides forsythus, Pr. intermedia, Prevotella nigrescens og Treponema denticola. Epstein-Barr virus er påvist i B-lymfocytter, og humant cytomegalovirus i monocytter/makrofager og T-lymfocytter fra parodontiet. Virusaktivering formodes at hæmme immunforsvaret og derved lette opformeringen af de patogene bakterier. Hypotesen er fremsat på basis af ganske få undersøgelser, hvor der er anvendt en særlig følsom PCR-teknik for at kunne detektere virus. Betydningen af tilstedeværelse af få virus er derfor langt fra afklaret.
Vi kan ikke udtale os sikkert om hvilke mikrobielle arter eller konsortier af arter der spiller en afgørende rolle i den parodontale destruktion. Den subgingivale mikrofloras umådelige kompleksitet er en forklaring. Vanskelighederne forbundet med at dyrke og identificere de subgingivale bakterier med specielle næringskrav er en anden. En trejde forklaring er at de patogene mikroorganismer tilhører den residente flora. De kan også påvises hos individer uden sygdom. Deres tilstedeværelse er ikke tilstrækkeligt bevis for at de forårsager eller deltager i infektionen. For endogene mikroorganismer må vi skelne mellem kolonisation og infektion. Kolonisation er en positiv interaktion mellem mikroorganismer og vært, som sker til gensidig fordel. Ved infektion er der en negativ påvirkning af mikroorganismerne på værtsorganismen. Som den økologiske plakhypotese beskriver, vil der ved parodontal destruktion være udviklet en mikrobiel sammensætning der overskrider tærsklen for et harmonisk forhold til værten. Balancen mellem mikrobiel påvirkning og værtsforsvaret kan beskrives ved flg. ligning:
Infektion = |
mikroflora: virulens x antal x tid |
immunforsvar: lokal/generelt |
Virulens
Virulens er en betegnelse for mikroorganismernes samlede evne til at fremkalde infektion. Den opdeles i evnen til at etablere sig, trænge ind og skade værten. For bakterier tilhørende den fastboende flora er det allerede vist at de kan etablere sig i mundhulen.
Klassiske eksempler på infektioner med invasion er tuberkulose, influenza og lungebetændelse, der alle er karakteriserede ved en primær kolonisering af slimhinder, hvorefter de patogene mikroorganismer invaderer og spredes i kroppen. Det er svært at påvise at der sker invasion ved parodontitis. Det var i mange år en stående diskussion hvorvidt der overhovedet sker invasion af parodontiet. Traumatiserende indgreb i pochen, fx depuration, pochedybdemåling og brug af tandtråd, skaber kortvarige bakteriæmier, fordi bakterier aktivt indføres i vævene ved processen. Den påviste forekomst af bakterier i de parodontale væv, som flere histologiske undersøgelser angiver at have vist, skyldes derfor ikke nødvendigvis en aktiv invasion af mikroorganismerne selv. De kan være indført ved sådanne procedurer. Et eksempel på selvstændig bakteriel invasion af parodontiet er akut, nekrotiserende, ulcerativ gingivitis, hvor det traumatiserede eller ulcererede epitel invaderes af bevægelige spirokæter, der kan vandre langt ind i bindevævet. Hvis de ikke rammes effektivt med antibiotica, leder det til de uhyggelige destruktioner ved noma. Bakteriel invasion af epitelceller som et primært skridt i infektionen er kun sparsomt undersøgt for orale bakterier. P. gingivalis og A. actinomycetemcomitans er nogle af de få undersøgte arter, og de kan invadere og formere sig i epitelceller in vitro, ligesom deres tilstedeværelse i orale epitelceller in vivo er påvist (12 – 14). Der sker formentlig invasion af toksiner og andre patogene produkter fra den subgingivale flora ind i de parodontale væv, men også dette område er nyt og ufuldstændigt belyst.
Evnen til at beskadige værten er åbenbar ved parodontitis. Den relative betydning af direkte vævsbeskadigelse forårsaget af mikroorganismer og indirekte beskadigelse forårsaget af inflammationsreaktionen er ikke fuldt klarlagt. Traditionelt opfatter vi inflammationsreaktionen som den vigtigste årsag til den parodontale nedbrydning. Men det er også tydeligt at der kan bestå et harmonisk forhold mellem mikroorganismer og vært uden parodontal nedbrydning i årevis på trods af inflammation (fx ved gingivitis der ikke udvikler sig til marginal parodontitis). Måske skal de vigtige, patogene mikroorganismer søges blandt dem der kan aktivere de vævsdestruerende dele af de inflammatoriske processer.
P. gingivalis, Pr. intermedia, B. forsythus og T. denticola er alle proteolytiske bakterier der kan nedbryde immunglobuliner og komplementfaktorer og således afvæbne immunforsvaret. P. gingivalis er især velundersøgt. Den producerer store mængder af adskillige proteolytiske enzymer hvormed den kan angribe stort set alle tænkelige værtsproteiner (15). Invaderende patogene mikroorganismer mødes altid med komplementaktivering, der kan føre til lyse af mikroorganismer eller fagocytose med polymorfnukleære leukocytter. P. gingivalis nedbryder komplementfaktorer, så denne proces hæmmes, og den unddrager sig også det specifikke immunforsvar ved at nedbryde antistoffer. Herved sætter P. gingivalis immunforsvaret ud af spillet og sikrer formentlig både sin egen og andre tilstedeværende bakteriers overlevelse. Desuden nedbryder den værtens egne proteaseinhibitorer, som indgår i reguleringen af inflammationsreaktionen. Dette er blot nogle af effekterne af P. gingivalis’ brede vifte af proteolytiske enzymer. Fælles for virkningerne er at de dysregulerer de normale immunforsvarsmekanismer og dermed formentlig bidrager til at ødelægge balancen mellem mikroorganismer og vært, så destruktion af parodontiet bliver resultatet. Måske er netop dette et eksempel på en nøglemekanisme der formår at ødelægge balancen mellem mikroorganismer og vært i parodontiet.
Antal
At den infektiøse dosis af patogene mikroorganismer varierer meget, kender man fra mange klassiske infektionssygdomme. Shigella kan give infektion med færre end 100 celler, mens Salmonella og Campylobacter kræver 104 – 105 for at forårsage sygdom. Også ved de parodontale sygdomme kræves der en vis load af mikroorganismer. Den skandinaviske skoles succes med at anvende total plakkontrol i behandling og profylakse af parodontitis bygger netop på at man ved at holde det totale antal mikroorganismer på et lavt niveau også reducerer risikoen for at nå en patogen kombination.
Tid
De fleste infektionssygdomme har relativt korte inkubationstider. Ved endogene sygdomme er ekspositionstiden uendelig. Der er derfor stor risiko for at de andre faktorer i infektionsligningen før eller senere spiller sammen, så der opstår sygdom. Derfor får så mange mennesker parodontitis.
Lokalt/generelt forsvar
Kroppens reaktion på en klassisk infektion er den uspecifikke inflammationsreaktion, klinisk sygdom og produktion af høje titre af specifikke antistoffer mod de patogene mikroorganismer. Det er helt indlysende at der foregår en inflammation i de parodontale væv som en reaktion på tilstedeværelsen af den subgingivale plak. Den indtræffer ved for høj bakteriebelastning i for lang tid. Reaktionen er uspecifik derved at den foregår på samme måde, uanset hvilke mikroorganismer der initierer den, og fortæller derfor ikke hvilke mikroorganismer der er betydningsfulde.
Et højt niveau af specifikke antistoffer er en sædvanlig reaktion på inficerende mikroorganismer ved klassiske infektionssygdomme. Ved en lokal aggressiv parodontitis (tidligere lokaliseret juvenil parodontitis (LJP)) kan der måles et højt antistofniveau mod A. actinomycetemcomitans og især mod dens leukotoksin. Det viser at netop denne form for parodontitis ligner de klassiske infektioner (16). Dette gælder for børn og unge. Hos voksne derimod er der ikke en tydelig sammenhæng mellem specifikke antistoffer, kolonisation med bestemte bakteriearter og parodontal sygdom. Snarere minder de relativt lave titre af antistoffer om de reaktioner man kan måle mod normalfloraen til enhver tid.
De parodontale sygdomme er inflammationsbetingede, dvs. vævsnedbrydningen skyldes primært immunforsvarets reaktion på den mikrobielle infektion. Inflammationsreaktionen er potentielt vævsdestruerende og reguleres af en lang række inhibitorer og regulative molekyler for ikke at løbe løbsk. Det er muligt at have en balance mellem den mikrobielle påvirkning og immunforsvaret, eksemplificeret ved patienter der i årevis har gingivitis, men ikke udvikler parodontitis. Samme patient kan have sites der forbliver inflammerede, men uden tab af fæste, og sites hvor balancen ikke opretholdes, og hvor der sker progredierende destruktion. If. infektionsligningen kan dette skyldes enhver af de medvirkende faktorer. Det er nærliggende at antage at der i en progredierende poche er en specifik sammensætning af floraen som immunforsvaret ikke kan hamle op med. Hvorfor kommer immunforsvaret til kort? Som eksemplificeret tidligere med P. gingivalis kan det skyldes at de specifikke mikroorganismer i denne poche kan unddrage sig visse dele af immunforsvaret og ligefrem dysregulere de normale immunforsvarsmekanismer, så inflammationsreaktionen kommer ud af kontrol.
Hos patienter med voldsom destruktion i hele parodontiet kan der være tale om en specifik kombination af særligt virulente mikroorganismer i alle pocher. Men der kan også være tale om systemiske defekter i immunsystemet. Pochens vigtigste komponenter for at forhindre infektion er granulocytter, antistoffer og komplement. Deres effektivitet bygger på samarbejde, hvor både antistoffer og komplement fungerer som forstærkningsmekanismer gennem opsonisering af bakterier for bedre kemotaksi og fagocytose med granulocytterne. Sygdomme som indebærer defekter i eller dårlig funktion hos granulocytter, fx diabetes mellitus, agranulocytose, neutropeni og evt. rygning, indebærer at ligevægten forskydes, og at lavere virulens, mindre dosis og kortere tid behøves, før den parodontale infektion indtræffer. Andre systemiske sygdomme og defekter med konsekvens for immunforsvaret, påvirkning fra medicin eller andet (fx rygning) samt lokale faktorer som dårlig drænage fremmer den parodontale infektion.
A. actinomycetemcomitans og lokal, aggressiv parodontitis (tidl. LJP)
Molekylærbiologiske undersøgelser har vist at der eksisterer en betydelig variation inden for de enkelte bakteriearter (17). For en del orale arter har det vist sig at næsten hver person har sin egen variant eller type.
A. actinomycetemcomitans (Fig. 2) er i mange undersøgelser kædet sammen med lokal, aggressiv parodontitis. Den producerer leukotoksin, der dræber leukocytter, og som anses for en vigtig virulensfaktor hos bakterien, fordi den derved kan undgå granulocytmedieret drab. Epidemiologiske undersøgelser af A. actinomycetemcomitans har vist at der eksisterer en selvstændig udviklingslinje, en klon, inden for arten, der producerer 10 – 20 gange mere af virulensfaktoren leukotoksin end andre isolater af A.actinomycetemcomitans. Den er altid associeret med tidlig og meget hurtigt forløbende parodontal destruktion, som ofte grupperer sig i familier og er terapiresistent. Klonen findes udelukkende hos personer af afrikansk afstamning (18), og den findes ikke hos sunde individer. Den er det hidtil klareste bud på en specifik parodontal sygdom hos individer af afrikansk afstamning. Hvorfor denne klon af A.actinomycetemcomitans ikke koloniserer hvide, vides endnu ikke. Associationen mellem A. actinomycetemcomitans og lokal, aggressiv parodontitis hos hvide er mindre klar. Det kunne måske skyldes at mængden af A. actinomycetemcomitans hos hvide er langt mere afgørende pga. den mindre leukotoksinproduktion hos de typer af A. actinomycetemcomitans der koloniserer hvide.
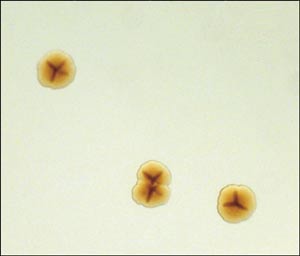
Fig. 2. Kolonier af Actinobacillus actinomycetemcomitans på agarplader. Bemærk den typiske stjernestruktur, som ses i koloniens indre ved gennemfaldende lys.
Den parodontale infektion
Af ovenstående fremgår at det i høj grad er antallet af mikroorganismer og deres aktivitet, der er afgørende for om der sker parodontal destruktion, samt at større mikrobiel aktivitet vil øge sandsynligheden for hurtigere udvikling af infektion. Mikroorganismernes aktivitet er ligeledes interessant i relation til den parodontologiske nomenklatur, der efterhånden kun skelner mellem to former for marginal parodontitis: aggressiv og kronisk parodontitis. Dette passer sammen med de to klassiske former for infektion: akut/subakut infektion og kronisk infektion. Den akutte/subakutte form (aggressiv parodontitis) kendetegnes ved høj aktivitet og hurtig tilvækst af mikroorganismer. Værten reagerer hurtigt, men uspecifikt med kraftig udtræden af leukocytter, især granulocytter, og i svære tilfælde abscesdannelse. Processen er karakteriseret ved vævsdestruktion, men det tager et stykke tid og er ikke synligt radiologisk før efter et par dage. Processen kan give almene symptomer, lymfeknuderne er ofte involverede, og der er risiko for generel spredning. Klinisk ser vi dette som en parodontal absces, mens mindre dramatiske, mere subakutte former er sværere at opdage. En parodontal destruktion der opstår på relativt kort tid, er ensbetydende med aggressiv parodontitis.
Den kroniske form (kronisk parodontitis) kendetegnes ved tilstedeværelse af mikroorganismer med relativt lav aktivitet. Værtsorganismen svarer med inflammation og udtræden af leukocytter, men her er lymfocytter de dominerende celler. Inflammationen medfører typisk en omdannelse af væv (ikke destruktion) til granulationsvæv, som er mere velegnet til at forhindre invasion af mikroorganismer og/eller deres virulensfaktorer. Der skabes en balance mellem værtsorganismen og mikroorganismerne med få symptomer og begrænset risiko for spredning. Prisen for denne balance er dog en vævsomdannelse med tab af det normale væv (parodontalligament og knogle) til fordel for granulationsvæv og nedvækst af epitel. På længere sigt giver den kroniske proces altså fæstetab.
English summary
Periodontal infection
The etiology of periodontal disease has been widely discussed during the last century. Periodontitis has been regarded as a non-specific, endogenous disease as well as a monospecific, exogenous disease. Both of these conceptions, however, contain shortcomings in their explanations of the distribution of periodontal destruction in the oral cavity. Presently, the dominating understanding of the development of periodontal disease is expressed in the ecological plaque hypothesis, which implies that periodontitis is caused by resident flora which may contain pathogenic species of bacteria. As microbiological methods of identification, and especially the application of molecular biological techniques, have been improved, the complexity of the subgingival plaque has increased, and even viruses have been detected. Whether or not periodontal destruction occurs is determined by the balance between, on the one hand, the composition and the virulence of the microflora and the number of bacteria in the gingival plaque and, on the other hand, the efficiency of the immune system, including both local and systemic factors. Further, whether aggressive or chronic periodontitis develops is influenced by the activity of the microflora.
Litteratur
1. Lövdal A, Arno A, Waerhaug J. Incidence of clinical manifestations of periodontal disease in light of oral hygiene and calculus formation. J Am Dent Assoc 1958; 56: 21 – 33.
2. Schei O, Waerhaug J, Lövdal A, Arno A. Alveolar bone loss as related to oral hygiene and age. J Periodontol 1959; 30: 7 – 16.
3. Lindhe J, Hamp S-E, Löe H. Plaque induced periodontal disease in beagle dogs. 4-year clinical, roentgenographical and histometrical study. J Periodontal Res 1975; 10: 243 – 55.
4. Goodson JM, Tanner ACR, Haffajee AD, Sornberger C, Socransky SS. Patterns of progression and regression of advanced destructive periodontal disease. J Clin Periodontol 1982; 9: 472 – 81.
5. Slots J, Ting M. Actinobacillus actinomycetemcomitans and Porphyromonas gingivalis in human periodontal disease: occurence and treatment. Periodontol 2000 1999; 20: 82 – 121.
6. Moore WEC, Moore LVH. The bacteria of periodontal diseases. Periodontol 2000 1994; 5: 66 – 77.
7. Socransky SS, Haffajee AD. Evidence of bacterial etiology: a historical perspective. Periodontol 2000 1994; 5: 7 – 25.
8. Marsh PD. Microbial ecology of dental plaque and its significance in health and disease. Adv Dent Res 1994; 8: 263 – 71.
9. Kroes I, Lepp PW, Relman DA. Bacterial diversity within the human subgingival crevice. PNAS 1999; 96: 14547 – 52.
10. Paster BJ, Boches SK, Galvin JL, Ericson RE, Lau CN, Levanos VA, Sahasrabudhe A, Dewhirst F. Bacterial diversity in human subgingival plaque. J Bacteriol 2001; 183: 3770 – 83.
11. Contreras A, Slots J. Herpesviruses in human periodontal disease. J Periodontal Res 2000; 35: 3 – 16.
12. Meyer DH, Mintz KP, Fives-Taylor PM. Models of invasion of enteric and periodontal pathogens into epithelial cells: a comparative analysis. Crit Rev Oral Biol Med 1997; 8: 389 – 409.
13. Madianos PN, Papapanou PN, Nannmark U, Dahlén G, Sandros J. Porphyromonas gingivalis FDC 381 multiplies and persists within human oral epithelial cells in vitro. Infect Immun 1996; 64: 660 – 4.
14. Rudney JD, Chen R, Sedgewick GJ. Intracellular Actinobacillus actinomycetemcomitans and Porphyromonas gingivalis in buccal epithelial cells collected from human subjects. Infect Immun 2001; 69: 2700 – 7.
15. Travis J, Banbuna A, Potempa J. The role of bacterial and host proteinases in periodontal disease. Adv Exp Med Biol 2000; 477: 455 – 65.
16. Ebersole JL, Taubman MA, Smith DJ, Genco RJ, Frey DE. Human antibody response to oral microorganisms I. Association of localized juvenile periodontitis with serum antibody response to Actinobacillus actinomycetemcomitans. Clin Exp Invest 1981; 47: 43 – 52.
17. Kilian M. Clonal basis of bacterial virulence. In: Guggenheim B, Shapiro S, editors. Oral biology at the turn of the century. Basel: Karger; 1998. p. 131 – 42.
18. Haubek D, Dirienzo JM, Tinoco EMB, Westergaard J, López NJ, Chung C-P, et al. Racial tropism of a highly toxic clone of Actinobacillus actinomycetemcomitans associated with juvenile periodontitis. J Clin Microbiol 1997; 35: 3037 – 42.
Søkeord for nettversjon: www.tannlegetidende.no: Gingivitt; Mikrobiologi; Periodontitt, marginal; Plakk
Korrespondance: Ellen V.G. Frandsen, Afdeling for Oral Biologi, Tandlægeskolen, Vennelyst Boulevard, Århus C, Danmark. E-post: ef@microbiology.au.dk
Artikkelen er fagfellevurdert.
Artikkelen siteres som:
Frandsen EV, Larsen T, Dahlén G. Den parodontale infektion. Nor Tannlegeforen Tid. 2004;114:30–4. doi:10.56373/2004-1-7